Morphological Description
Life History
Distribution
Habitat
Roost Sites and Roosting Patterns
Emergence and Flight Pattern
Foraging Behaviour
Echolocation Calls
Status and Protection |
|
|
Morphological Description
- Dorsal fur is golden or reddish-brown. Ventral fur is paler and duller.
- Juveniles are darker and duller than adults.
- If disturbed during torpor may feign death. Emits a musky smell when irritated.
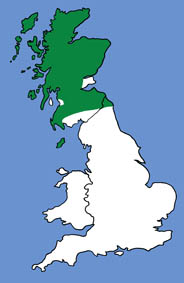
- Noctules have a post-calcarial lobe and a mushroom-shaped tragus. They can be distinguished from Leisler's bats by the length of the forearm. The forearm of noctule bats is longer than 47mm.
- Adult males have a long, slender penis.
- Average weight (as given by Greenaway & Hutson, 1990) 18-40 g.
The diagram below gives important average body measurements for noctule bats (Greenaway & Hutson, 1990).
|
Back to top |
Life History
- Mate from August through to October.
- Male and female bats can store sperm for up to seven months without the sperm losing their fertilising capacity (Racey, 1991).
- Gestation period is 70-73 days with a single offspring being born in June or July (Racey, 1991).
- Maximum age recorded in Europe is 12 years (Schober & Grimmberger, 1989).
Back to top
|
Distribution |
|
|
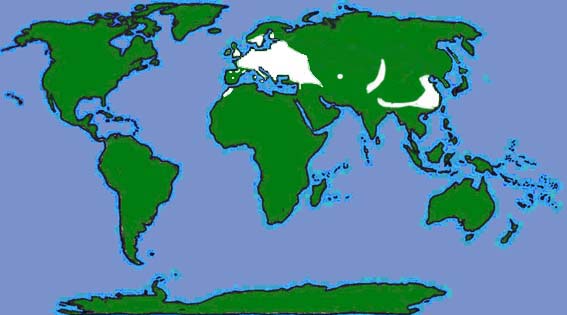 |
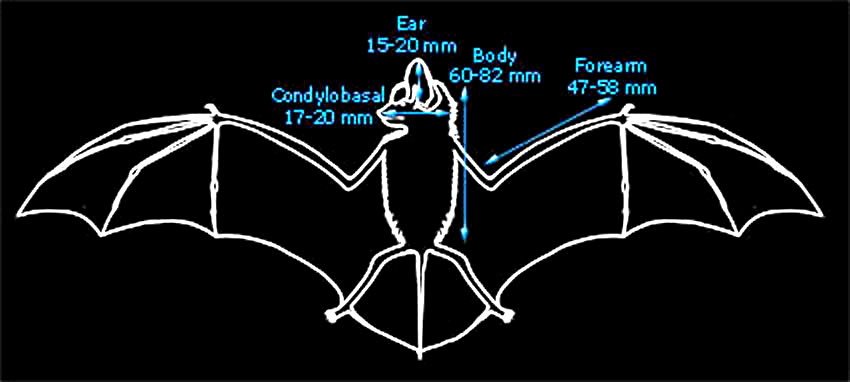
| The British and World distributions are shown by the white areas of the maps above (as given by Richardson, 2000 and Corbet & Harris, 1991 respectively). |
|
Back to top
|
Habitat |
|
- Noctules prefer open habitats and rivers or lakes (Vaughan et al., 1997).
- More often found in lowland areas and those with old forests, rivers and marshland.
- Uncommon in cities.
- The photograph on the left shows a typical habitat of noctules.
|
| Roost Sites and Patterns |
- Summer roosts: Females are found in nursery roosts from March. Males are solitary or live in small, social groups during the summer. Roosts are found in woodpecker holes and natural cavities in mature, deciduous trees, especially beech (Racey, 1991).
- Females move between territorial mating roosts which are established by males in autumn using vocalisations. Several females may occupy the roost at any one time.
- Kronwitter (1988) found that during the nursing period male and female noctules roost in distinctly separate geographical areas. He also observed frequent roost changing behaviour, which may be for hygiene reasons or possibly to search for new foraging areas.
- Winter roosts: large, mixed sex colonies are found in tree holes, deep rock crevices or buildings.
- Boonman (2000) investigated roost site choice by noctules. Noctules prefer woodpecker cavities to natural tree cavities and so may be dependent on the presence of woodpeckers in their habitats. They also prefer to roost near forest edges, perhaps to limit energy expenditure in flying to their feeding site.
- Summer and winter roosts sites are often miles apart.
- Use bat boxes more readily than other species.
Back to top
|
Emergence and Flight Pattern |
- Often the first species to appear in the evening, emerging 0-40 minutes after sunset (Racey, 1991; Russ, 1999) and occasionally before sunset.
- Median emergence time is 5 minutes after sunset (Jones & Rydell, 1994).
- Jones (1995) found that noctules emerged earlier when females were lactating, potentially increasing their risk of predation when energetic demands are high.
- Flies high and fast.
- Flight is straight but includes deep dives to chase insects and occasional glides.
- Not a very agile flier and hence flies over open habitats.
Back to top
|
Foraging Behaviour |
- Foraging activity peaks for one hour at dusk and half an hour at dawn.
- The emergence time and length of foraging activity of noctules are influenced by temperature, precipitation and moonlight. The bats reduced intraspecific competition by foraging at different times of the night to their roostmates. Noctules forage exclusively in open habitats; the bats' wing morphology and echolocation calls are well adapted for this type of foraging (Kronwitter, 1998).
- The diet of noctules mainly consists of Diptera, with both crepuscular and diurnal species being eaten (Vaughan, 1997). Lepidoptera and Coleoptera are also important dietary items. Aerial hawking is the predominant foraging strategy used.
- Prey is caught and consumed during flight. May also forage around lights or in the substrate.
|
 |
|
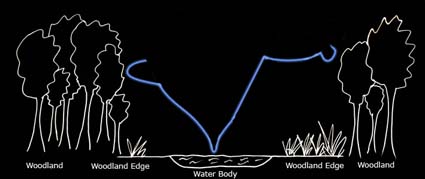
Marked in blue on the diagram above is a typical foraging path of noctules (based on Russ, 1999).
Back to top
|
Echolocation Calls |
|
|
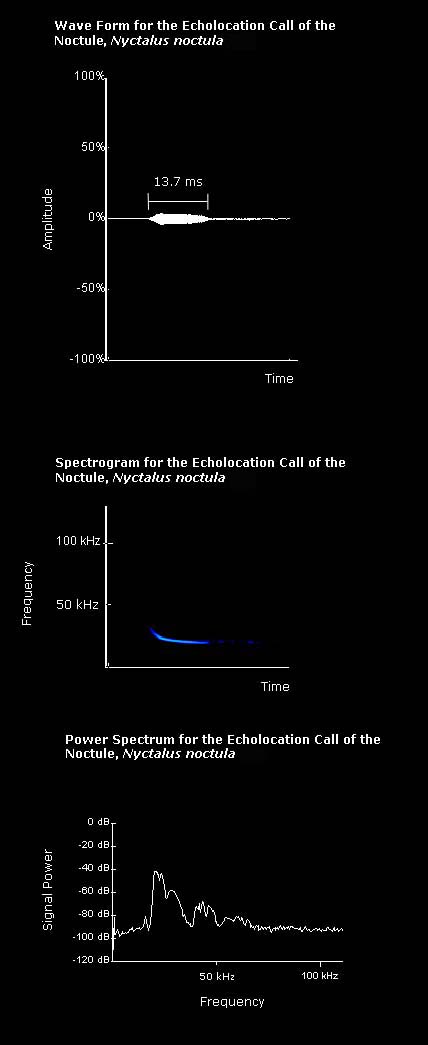 |
The echolocation call of noctules has a narrow bandwidth. |
|
To listen to the call of the noctule bat click here
Size of sound file: 34.4 KB
|

|
| For details of how the echolocation calls were recorded click here. |
|
Average values for a noctule echolocation call, as given by Vaughan et al. (1997), are listed below:
Interpulse interval: 336ms
Call duration: 19.6ms
Minimum frequency: 18.2kHz
Maximum frequency: 20.3kHz
The power spectrum on the left shows that the maximum power of the call is at a frequency of approximately 21kHz.
|
Back to top
|
|
| Status and Protection
- The British pre-breeding population was estimated at 50,000 in 1995 (45,000 in England, 250 in Scotland and 4750 in Wales) (Harris et al., 1995).
- Have become very rare in Britain, particularly in agricultural areas.
- Noctules are at low risk of extinction worldwide (IUCN status, 2001).
- Destruction of roost sites through deforestation and removal of winter roosts in buildings threatens remaining populations.
- Bat boxes can be used to help maintain existing populations.
Back to top
|
 © School of Biological Sciences, University of Bristol 2005. Last modified 24th February 2005. © School of Biological Sciences, University of Bristol 2005. Last modified 24th February 2005. |